Origin of Life Theories: Metabolism-first vs. Replicator-first Hypotheses

Introduction
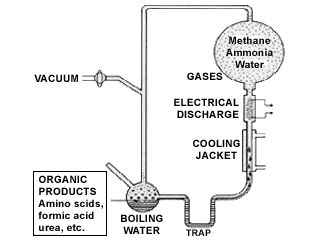
Origin of life research tends to fall into either of two camps. The first experiments done by Stanley Miller in 1953 demonstrated that mixtures of reducing gases, thought to be present in the primordial earth, when subjected to electrical discharges, produced many organic compounds, including several amino acids. Such experiments suggested that the components of living organisms might have arisen naturalistically on earth. Subsequent research revealed that Miller used the wrong gases, but origin of life research was off and running. When researchers ran into the problem of how these cellular building blocks might be organized, it was suggested that a replicator was first required. However, since DNA (the current reigning replicator) requires an extensive array of protein components in order to replicate, it was hypothesized that RNA coudl catalyze its own replication, resulting in the "RNA world" hypothesis.
Replicator-first problems
In 2007, Robert Shapiro wrote a scathing critique of the RNA world hypothesis for Scientific American.1 Shapiro pointed out that Miller's spark experiment produced amino acids and tars, but no nucleoside RNA bases. Scientists soon were using purified chemicals in highly contrived conditions, with Shapiro noting that "kilograms of starting material were required to produce milligrams of product." Shapiro also indicated that some experiments required that a large lagoon be dried to the size of a puddle without the loss of any starting material, even though no such features exist on earth today. In addition, most reactions produce products that are inherently unstable, decomposing within a short period of time to unusable products. Because of the enormous problems associated with the spontaneous synthesis of RNA, some researchers have opted for a pre-RNA world, in which smaller molecules substitute for RNA. However, none of the proposed compounds have ever been shown to be able to catalyze their own synthesis. In addition, numerous spontaneously-produced inhibitors block pre-biotic chemistry, requiring the use of purified compounds. Shapiro concluded that "The majority of origin-of-life scientists who still support the RNA-first theory either accept this concept (implicitly, if not explicitly) or feel that the immensely unfavorable odds were simply overcome by good luck."
More examples of why replicator-first scenarios are unlikely can be found below.
Metabolism-first problems
Shapiro's critique of replicator-first hypotheses included a discussion of how a metabolism-first mechanism might have produced the first living organisms. Although some of the starting materials are simpler, the process requires many more steps, including:
- A boundary - some kind of primitive membrane to separate living from non-living components.
- An energy source - usually some kind of oxidation-reduction system, which could be organic or mineral-based.
- A coupling mechanism - so that energy can be used for a useful purpose or in reproduction.
- A chemical network - some kind of metabolic pathway.
- The network must grow and reproduce. - at some point a replicator needs to be invented.
Leslie Orgel's rebuttal
Shapiro's description contained a wealth of generalities, but no specifics or a discussion of the numerous problems associated with metabolism-first origin of life scenarios. During 2007, until his death in October, Leslie Orgel wrote a rebuttal to the metabolism-first hypothesis, published posthumously in 2008.2 In his article, Orgel examined various metabolic pathways to determine if it were feasible that such pathways could have operated in pre-biotic environments.
Carbon fixation
In order for living organisms to exist, they must be able to fix carbon from the environment into organic molecules. For the reverse citric acid cycle, although no specific mineral catalytic agents have been identified, it is possible that such catalysts might exist. However, specific reactions of the cycle would be problematic. The uncatalyzed reaction that converts malic acid to fumaric acid are extremely slow, with a half-life of 10,000 years. The reaction of pyruvic acid and α-ketoglutaric acid to produce oxaloacetic acid is energy requiring, so that the reverse reaction is thermodynamically favored. So, an additional energy-producing reaction would need to be coupled to the reaction. However, a possible coupling of the hydrolysis of an inorganic pyrophosphate or polyphosphate would require an additional catalyst or else the introduction of phosphate groups globally would destroy the cycle. Reactions producing pyruvic acid from acetic acid and α-ketoglutaric acid from succinic acid are extremely unfavorable using carbon dioxide as the carbon source. Orgel suggested that carbon monoxide might make a better carbon source, without acknowledging that this source would be very limited on the prebiotic earth.
However, the main problem in running the reverse citric acid cycle is the existence of side reactions that would disrupt the cycle. The splitting of isocitric acid to succinic acid and glyoxylic acid, would return succinic acid to the cycle, reducing its efficiency because some of the acetate or oxaloacetate would be shunted to glyoxylic acid. Likewise, the splitting of malic acid to glyoxylic acid and acetic acid would return acetic acid to the cycle. Another side reaction involves reductive carboxylation reactions, which would be expected to accept malic acid instead of acetic and succinic acids, which would move reactants irreversibly out of the cycle. Another problem is that specific catalysts would be required for the reduction of oxaloacetic acid, oxalosuccinic acid, and fumaric acid, since non-specific catalysts would also reduce pyruvic acid and α-ketoglutaric acid, disrupting the cycle. Orgel comments, "It is likely that such catalysts could be constructed by a skilled synthetic chemist, but questionable that they could be found among naturally occurring minerals or prebiotic organic molecules." Carboxylation reactions would also require specific catalysts to prevent the diversion of acetate to malonate. Likewise, the citrate lyase reaction would require specific catalysts, since the conversion of a carboxylic acid group to a thioester would occur with nearly all molecules involved in the reverse citric acid cycle, leading to destructive side reactions. The requirement of multiple, specific mineral catalysts at exactly the same location seems implausible to Orgel, who says, "It is not completely impossible that sufficiently specific mineral catalysts exist for each of the reactions of the reverse citric acid cycle, but the chance of a full set of such catalysts occurring at a single locality on the primitive Earth in the absence of catalysts for disruptive side reactions seems remote in the extreme."
The three other carbon fixation pathways, the Calvin cycle, the hydroxypropionate cycle, and the reductive acetyl-CoA linear pathway, suffer from similar problems of disruptive side reactions in the absence of specific catalysts. Some scientists are so enamored with biological evolution that they propose that cycles of reactions can evolve independently of a replicator. According to Dr. Orgel, "The scheme should not make unreasonable demands on the efficiency and specificity of the various external and internally generated catalysts that are supposed to be involved. Without such a description, acceptance of the possibility of complex nonenzymatic cyclic organizations that are capable of evolution can only be based on faith, a notoriously dangerous route to scientific progress."
Peptide cycles
Stuart Kauffman has proposed that prebiotic amino acids self-assembled to form catalytic peptides that have auto-catalytic properties.3 In theory, these peptides would be self-reproducing and could produce a complete cycle that could be used to fix carbon. Some of the theory has been reproduced in the laboratory. Certain designed 32-mer peptides have been shown to catalyze the addition of amino acids. However, the problem is that there is no way to get peptides of that length through pre-biotic chemistry. Orgel says, "Ghadiri's experiments illustrate 'intelligent design' of input peptides, not spontaneous self-organization of polymerizing amino acids", with "intelligent design" being an obvious, though accurate, slam on what was actually shown experimentally. Unfortunately, there were no organic chemists present on the prebiotic earth, unless one wants to consider the ultimate Intelligent Designer. Orgel explained that the discrimination and catalytic abilities of 10-mer or smaller peptides would not be sufficient to produce any kind of self-reproducing cycle. Catalytic enzymes are 100-mer to 1000-mer in order to specify their remarkably-specific catalytic ability. Small peptides do not form the stable three-dimensional structures required to consistently maintain any kind of catalytic activity. Even most larger enzymes do not spontaneously form the proper three-dimensional structure, but must be specifically folded by specialized macromolecular machines.
Orgel's conclusions
Leslie Orgel ends his attack on metabolism-first hypotheses with an admission that replicator-first theories are deeply flawed, but that metabolism-first scenarios are no less flawed, but "must stand on their own":
"The most serious challenge to proponents of metabolic cycle theories—the problems presented by the lack of specificity of most nonenzymatic catalysts—has, in general, not been appreciated. If it has, it has been ignored. Theories of the origin of life based on metabolic cycles cannot be justified by the inadequacy of competing theories: they must stand on their own."
Orgel's last sentence is quite amusing:
"However, solutions offered by supporters of geneticist or metabolist scenarios that are dependent on 'if pigs could fly' hypothetical chemistry are unlikely to help."
Metabolist systems can't evolve
Vasas V, Szathm�ry E, Santos M. 2010. Lack of evolvability in self-sustaining autocatalytic networks constraints metabolism-first scenarios for the origin of life. Proc Natl Acad Sci U S A. 107:1470-1475.
Conclusion

The origin of life wars are in full swing as proponents of each hypothesis, replicator-first and metabolism-first theories, show how each other's theories cannot possibly work on the prebiotic earth. All current data indicates that the origin of life is extremely unlikely to have occurred through prebiotic chemistry on the early earth. It was this realization, in 1972 during my undergraduate studies at USC, that convinced me that there must have been an Intelligent Creator who made the first life on earth—changing my worldview from atheism to deism. Even after all those years, the evidence in favor of a naturalistic origin of life has not significantly improved.
Related Pages
- Is the Chemical Origin of Life (Abiogenesis) a Realistic Scenario?
- Origin of Life: Earth's Early Atmosphere Wasn't Reducing
- The Origin of Homochirality: A Major Problem for Origin of Life Theories
- Evolution Deception in California State High School Biology Textbook Biology: Principles & Explorations
- Problems with the Origin of Biological Membranes Under Prebiotic Conditions
- Cell Membrane-Like Organic Vesicles Formed in Conditions Mimicking Interstellar Clouds?
- What's Wrong With NASA's Arsenic-Eating Bacteria Study?
- NASA Scientist Discovers Alien Life in Meteorites - Again! NOT!
- Origin of life: latest theories/problems
- The Origin of Life on Planet Earth
- Book Review: Origins of Life: Biblical and Evolutionary Models Face Off
Related Materials
 Origins of Life:
Biblical and Evolutionary Models Face Off
Origins of Life:
Biblical and Evolutionary Models Face Off![]() by Fazale Rana and Hugh Ross. Probably the single most potent
scientific argument against atheism is the problem with a naturalistic origin of
life. This very problem led me to become a deist as a biology major at USC in
the early 1970's. The problems for atheists have gotten no better since that
time. In fact, the last 30+ years of research have turned up even more problems
than those that existed when I first studied the theories. Fuz Rana (a
biochemist) and Hugh Ross (an astrophysicist) have teamed up to write the
definitive up-to-date analysis of the origin of life. The book examines the origins of life from the
perspectives of chemistry, biochemistry, astronomy, and the Bible. A biblical
creation model is presented along side the naturalistic models to help the
reader decide which one fits the data better. This is an excellent book to give
to your unbelieving friends, since it presents a testable creation model that is
clearly superior to any naturalistic model.
by Fazale Rana and Hugh Ross. Probably the single most potent
scientific argument against atheism is the problem with a naturalistic origin of
life. This very problem led me to become a deist as a biology major at USC in
the early 1970's. The problems for atheists have gotten no better since that
time. In fact, the last 30+ years of research have turned up even more problems
than those that existed when I first studied the theories. Fuz Rana (a
biochemist) and Hugh Ross (an astrophysicist) have teamed up to write the
definitive up-to-date analysis of the origin of life. The book examines the origins of life from the
perspectives of chemistry, biochemistry, astronomy, and the Bible. A biblical
creation model is presented along side the naturalistic models to help the
reader decide which one fits the data better. This is an excellent book to give
to your unbelieving friends, since it presents a testable creation model that is
clearly superior to any naturalistic model.
References
- Shapiro, R. 2007. A Simpler Origin for Life. Scientific American February 12, 2007.
- Orgel, L. E. 2008. The Implausibility of Metabolic Cycles on the Prebiotic Earth. PLoS Biol. 6: e18.
- Kauffman S. A. 1986. Autocatalytic sets of proteins. J. Theor. Biol. 119: 1ñ24.
- Lee, D. H., J. R. Granja, J. A. Martinez, K. Severin and M. R. Ghadiri. 1996. A self-replicating peptide. Nature 382: 525-528.
http://www.godandscience.org/evolution/replicator_vs_metabolism.html
Last Modified February 25, 2010

